-es707.png)
1
f. Biol. La mitocondria es un orgánulo presente en el citoplasma de las células eucariotas en donde se produce la respiración celular (fosforilación oxidativa), generando la mayoría de los fosfatos de alta energía como el ATP.
Posee una forma esférica o alargada, y posee membrana interna y externa.
Las mitocondrias son la fuente de producción de energía dentro de las células eucarióticas. Son orgánulos semiautónomos y auto-reproductores, que residen en el citoplasma.
Convirtiendo los metabolitos de energía celular en el Ciclo de Krebs, a través del proceso de fosforilación oxidativa producen el trifosfato de adenosina (ATP), que se utiliza para potenciar otros procesos en la célula.
Su función es esencial para la producción eficiente de energía. Sin ellos, las células eucarióticas dependerían de la glicólisis anaeróbica para su ATP.
La glicólisis libera muy poca energía libre pero en las mitocondrias el metabolismo de los azúcares es mucho más eficiente y proporciona 15 veces más ATP que el que se produce a través de la glicólisis.
Las mitocondrias ocupan una gran parte del volumen citoplasmático de las células eucarióticas. Son varillas formadas por una membrana interior y otra exterior.
La membrana exterior limita el orgánulo.
La membrana interna se pliega sobre sí misma formando las cristas mitocondriales, dando la apariencia de particiones y cámaras dentro del organelo en sección transversal.
El número y la forma de las cristas varían según el tipo de tejido y el organismo. Las cristas sirven para aumentar la superficie de la membrana interior.
Las mitocondrias contienen su propio genoma, que está separado y es distinto del genoma de la célula.
Teóricamente, las mitocondrias pueden haber sido organismos unicelulares separados en un momento dado y fueron subsumidos en una relación simbiótica en las células eucarióticas en algún momento del proceso evolutivo.
La similitud de los genes entre el genoma mitocondrial y el genoma de Rickettsia prowazekii sugiere que esta bacteria es la más estrechamente relacionada con las mitocondrias hasta la fecha.
Normalmente se ven como cilindros inflexibles de aproximadamente 0,5-1 micrómetro, las mitocondrias se parecen a las bacterias.
Las mitocondrias son en realidad muy móviles y cambian de forma con frecuencia. Incluso pueden fusionarse con otras mitocondrias y luego separarse.
A menudo se asocian con microtúbulos en el citoplasma que determinan la orientación y distribución de las mitocondrias.
En algunas células forman largas cadenas móviles y en otros tipos de células permanecen fijas en una posición que proporciona ATP directamente a un sitio de alto consumo de ATP como las miofibrillas adyacentes del músculo cardíaco o el flagelo del esperma.
Las mitocondrias se vieron por primera vez en un microscopio de luz en el siglo XIX, pero no se entendieron realmente hasta que se desarrollaron procedimientos para aislar muestras intactas en 1948.
Las mitocondrias se suelen recuperar de células hepáticas purificadas que contienen de 1000 a 2000 mitocondrias cada una, ocupando alrededor de una quinta parte del volumen celular.
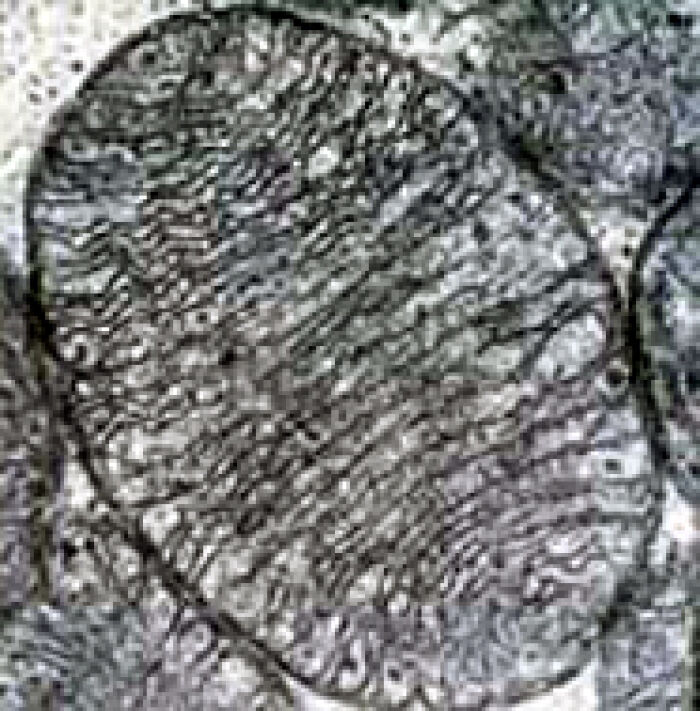
Micrografía electrónica de una sola mitocondria que muestra la disposición organizada de la matriz proteica y la membrana mitocondrial interna. CC
El piruvato formado en la glicólisis en la segunda etapa de la oxidación anaeróbica es transportado a las mitocondrias.
Las mitocondrias oxidan el piruvato con O2 a CO2 generando 34 de las 36 moléculas de ATP cuando la glucosa se convierte en CO2
Hay dos membranas estructurales y distintas y dos compartimentos o espacios internos. Las dos membranas están separadas por el espacio inter-membranas.
El espacio encerrado dentro de la membrana interna es la matriz, una región moderadamente densa con hebras de ADN, ribosomas o pequeños gránulos con los que las mitocondrias codifican una porción de sus propias proteínas.
La membrana externa de las mitocondrias contiene la proteína de transporte porina. La porina forma canales acuosos a través de la bicapa lipídica de las mitocondrias, que se asemeja a un tamiz permeable a través del cual las moléculas, incluyendo las pequeñas proteínas, de 5 kiloDaltons o menos pueden pasar al espacio inter-membrana.
La membrana interior, en comparación, es impermeable a la mayoría de las moléculas. En otras palabras, el espacio inter-membrana es químicamente como el citosol, mientras que la matriz interna de las mitocondrias contiene un selecto conjunto de moléculas.
La membrana interna constituye el marco para los procesos de transporte de electrones que producen la mayor parte del ATP de la célula.
Las enzimas incrustadas en la membrana mitocondrial interna son esenciales para el proceso de fosforilación oxidativa que genera la mayor parte del ATP de la célula animal.
Extremadamente enrevesado, su inflexión forma las crestas que se proyectan en la matriz del compartimiento interno de la mitocondria y aumentan significativamente el área de la membrana interna. Las cristas varían en volumen para los diferentes tipos de células.
El número de cristas en una mitocondria en una célula del miocardio, por ejemplo, es tres veces mayor que el de las mitocondrias en una célula del hígado. También hay una mayor demanda de ATP en las células del miocardio que en las del hígado.
En las procariotas, la membrana plasmática se utiliza para producir ATP. Sin embargo, las células eucarióticas reservan esta función para las membranas en organelos convertidores de energía, mitocondrias y plastidos y la membrana plasmática se reserva para otros procesos de transporte. Las mitocondrias y los plastidos son morfológicamente notables por sus extensas membranas internas.
La membrana interna es una bicapa lipídica que contiene una alta proporción de cardiolipina (difosfato-glicerol). Reduce suficientemente la permeabilidad de la membrana a los protones, haciendo la membrana especialmente impermeable a los iones y estableciendo las condiciones que permiten establecer una fuerza motriz de los protones a través de ella.
La membrana interna también contiene varias proteínas de transporte diferentes que la hacen selectivamente permeable a moléculas más pequeñas que son metabolizadas o utilizadas en las enzimas mitocondriales de la matriz.
La proteína constituye un porcentaje más alto de la membrana interna que en cualquier otra membrana de la célula, el 76%. La membrana interna contiene muchas partículas intramembrana ricas en proteínas, incluyendo los complejos F0F1 que sintetizan ATP y los que transportan electrones a O2 desde el NADH o el dinucleótido de adenina de flauta reducida (FADH2).
La membrana interna también contiene proteínas de transporte que pasan moléculas impermeables, como el ADP y el Pi, para pasar del citosol del espacio inter-membrana a través de la membrana interna a la matriz interna. Las proteínas también transportan otras moléculas en sentido inverso, como el ATP, desde la matriz al citosol.
Junto con la membrana interna, la matriz contenida en el compartimiento interno de las mitocondrias es el área funcional primaria de las mitocondrias.
La matriz incluye enzimas que metabolizan el piruvato y los ácidos grasos para producir acetil CoA junto con enzimas que oxidan el acetil CoA en el ciclo de Kreb. El ciclo de Kreb produce, entre otras cosas, NDAH que es la fuente primaria de electrones transportados a través de la cadena respiratoria. Las enzimas de la cadena respiratoria están incrustadas en la membrana mitocondrial interna, esencial allí para el proceso de fosforilación oxidativa.
En un proceso similar a la replicación en las células bacterianas, cuando una mitocondria alcanza un cierto tamaño, sufren una fisión, el surco de la membrana interna y externa que las pellizca en dos mitocondrias hijas. Antes de la fisión se replican su ADN.
Las mitocondrias al final de su vida son eliminadas por medio de la autofagia. El retículo endoplasmático celular se envuelve alrededor de la mitocondria formando una vacuola. Las vesículas del complejo de Golgi que contienen hidrolasas se unen entonces a la vacuola autofágica que luego degrada el contenido de la vacuola, la mitocondria.
El uniportador mitocondrial en la membrana interna mueve el Ca2+ hacia la matriz que a su vez inicia la activación de las deshidrogenasas en el ciclo del ácido cítrico, las concentraciones celulares de Ca2+ esencialmente desencadenando variaciones en la actividad de la mitocondria.
Esto establece un claro vínculo entre el metabolismo de la energía y la actividad celular que implica una relación funcional con el retículo endoplasmático.
La absorción de Ca2+ es aparentemente rápida, mientras que el Ca2+ se libera de nuevo en la célula durante un período más largo, lo que indica que las mitocondrias actúan como un amortiguador. Esta función de amortiguación tiene un impacto en los potenciales terminales presinápticos y puede estar implicada en la propagación de las ondas citosólicas de Ca2+.
La sobrecarga de las mitocondrias con Ca2+ puede estar relacionada con la apoptosis neuronal (muerte celular) a través de dos posibles resultados:
a) La disminución o pérdida del gradiente de protones de la membrana mitocondrial interna, que lleva a la terminación de la producción de ATP;
b) Pueden liberarse sustancias proactivas como el citocromo c en el protoplasma de la célula. Estas sustancias proactivas a su vez activarán caspasas que darán lugar a necrosis o apoptosis según el grado de pérdida de producción de ATP y la supervivencia de un número suficiente de mitocondrias.
Posee una forma esférica o alargada, y posee membrana interna y externa.
-es.svg){kind=link}
Funciones de las mitocondrias
Las mitocondrias son la fuente de producción de energía dentro de las células eucarióticas. Son orgánulos semiautónomos y auto-reproductores, que residen en el citoplasma.
Convirtiendo los metabolitos de energía celular en el Ciclo de Krebs, a través del proceso de fosforilación oxidativa producen el trifosfato de adenosina (ATP), que se utiliza para potenciar otros procesos en la célula.
Su función es esencial para la producción eficiente de energía. Sin ellos, las células eucarióticas dependerían de la glicólisis anaeróbica para su ATP.
La glicólisis libera muy poca energía libre pero en las mitocondrias el metabolismo de los azúcares es mucho más eficiente y proporciona 15 veces más ATP que el que se produce a través de la glicólisis.
Las mitocondrias ocupan una gran parte del volumen citoplasmático de las células eucarióticas. Son varillas formadas por una membrana interior y otra exterior.
La membrana exterior limita el orgánulo.
La membrana interna se pliega sobre sí misma formando las cristas mitocondriales, dando la apariencia de particiones y cámaras dentro del organelo en sección transversal.
El número y la forma de las cristas varían según el tipo de tejido y el organismo. Las cristas sirven para aumentar la superficie de la membrana interior.
Las mitocondrias contienen su propio genoma, que está separado y es distinto del genoma de la célula.
Teóricamente, las mitocondrias pueden haber sido organismos unicelulares separados en un momento dado y fueron subsumidos en una relación simbiótica en las células eucarióticas en algún momento del proceso evolutivo.
La similitud de los genes entre el genoma mitocondrial y el genoma de Rickettsia prowazekii sugiere que esta bacteria es la más estrechamente relacionada con las mitocondrias hasta la fecha.
Características generales
Normalmente se ven como cilindros inflexibles de aproximadamente 0,5-1 micrómetro, las mitocondrias se parecen a las bacterias.
Las mitocondrias son en realidad muy móviles y cambian de forma con frecuencia. Incluso pueden fusionarse con otras mitocondrias y luego separarse.
A menudo se asocian con microtúbulos en el citoplasma que determinan la orientación y distribución de las mitocondrias.
En algunas células forman largas cadenas móviles y en otros tipos de células permanecen fijas en una posición que proporciona ATP directamente a un sitio de alto consumo de ATP como las miofibrillas adyacentes del músculo cardíaco o el flagelo del esperma.
Las mitocondrias se vieron por primera vez en un microscopio de luz en el siglo XIX, pero no se entendieron realmente hasta que se desarrollaron procedimientos para aislar muestras intactas en 1948.
Las mitocondrias se suelen recuperar de células hepáticas purificadas que contienen de 1000 a 2000 mitocondrias cada una, ocupando alrededor de una quinta parte del volumen celular.
Micrografía electrónica de una sola mitocondria que muestra la disposición organizada de la matriz proteica y la membrana mitocondrial interna. CC
{kind=link}
Función
El piruvato formado en la glicólisis en la segunda etapa de la oxidación anaeróbica es transportado a las mitocondrias.
Las mitocondrias oxidan el piruvato con O2 a CO2 generando 34 de las 36 moléculas de ATP cuando la glucosa se convierte en CO2
Subestructura
Hay dos membranas estructurales y distintas y dos compartimentos o espacios internos. Las dos membranas están separadas por el espacio inter-membranas.
El espacio encerrado dentro de la membrana interna es la matriz, una región moderadamente densa con hebras de ADN, ribosomas o pequeños gránulos con los que las mitocondrias codifican una porción de sus propias proteínas.
Membrana exterior
La membrana externa de las mitocondrias contiene la proteína de transporte porina. La porina forma canales acuosos a través de la bicapa lipídica de las mitocondrias, que se asemeja a un tamiz permeable a través del cual las moléculas, incluyendo las pequeñas proteínas, de 5 kiloDaltons o menos pueden pasar al espacio inter-membrana.
La membrana interior, en comparación, es impermeable a la mayoría de las moléculas. En otras palabras, el espacio inter-membrana es químicamente como el citosol, mientras que la matriz interna de las mitocondrias contiene un selecto conjunto de moléculas.
Membrana interna
La membrana interna constituye el marco para los procesos de transporte de electrones que producen la mayor parte del ATP de la célula.
Las enzimas incrustadas en la membrana mitocondrial interna son esenciales para el proceso de fosforilación oxidativa que genera la mayor parte del ATP de la célula animal.
Extremadamente enrevesado, su inflexión forma las crestas que se proyectan en la matriz del compartimiento interno de la mitocondria y aumentan significativamente el área de la membrana interna. Las cristas varían en volumen para los diferentes tipos de células.
El número de cristas en una mitocondria en una célula del miocardio, por ejemplo, es tres veces mayor que el de las mitocondrias en una célula del hígado. También hay una mayor demanda de ATP en las células del miocardio que en las del hígado.
En las procariotas, la membrana plasmática se utiliza para producir ATP. Sin embargo, las células eucarióticas reservan esta función para las membranas en organelos convertidores de energía, mitocondrias y plastidos y la membrana plasmática se reserva para otros procesos de transporte. Las mitocondrias y los plastidos son morfológicamente notables por sus extensas membranas internas.
La membrana interna es una bicapa lipídica que contiene una alta proporción de cardiolipina (difosfato-glicerol). Reduce suficientemente la permeabilidad de la membrana a los protones, haciendo la membrana especialmente impermeable a los iones y estableciendo las condiciones que permiten establecer una fuerza motriz de los protones a través de ella.
La membrana interna también contiene varias proteínas de transporte diferentes que la hacen selectivamente permeable a moléculas más pequeñas que son metabolizadas o utilizadas en las enzimas mitocondriales de la matriz.
La proteína constituye un porcentaje más alto de la membrana interna que en cualquier otra membrana de la célula, el 76%. La membrana interna contiene muchas partículas intramembrana ricas en proteínas, incluyendo los complejos F0F1 que sintetizan ATP y los que transportan electrones a O2 desde el NADH o el dinucleótido de adenina de flauta reducida (FADH2).
La membrana interna también contiene proteínas de transporte que pasan moléculas impermeables, como el ADP y el Pi, para pasar del citosol del espacio inter-membrana a través de la membrana interna a la matriz interna. Las proteínas también transportan otras moléculas en sentido inverso, como el ATP, desde la matriz al citosol.
Matriz
Junto con la membrana interna, la matriz contenida en el compartimiento interno de las mitocondrias es el área funcional primaria de las mitocondrias.
La matriz incluye enzimas que metabolizan el piruvato y los ácidos grasos para producir acetil CoA junto con enzimas que oxidan el acetil CoA en el ciclo de Kreb. El ciclo de Kreb produce, entre otras cosas, NDAH que es la fuente primaria de electrones transportados a través de la cadena respiratoria. Las enzimas de la cadena respiratoria están incrustadas en la membrana mitocondrial interna, esencial allí para el proceso de fosforilación oxidativa.
Ciclo de vida
Replicación
En un proceso similar a la replicación en las células bacterianas, cuando una mitocondria alcanza un cierto tamaño, sufren una fisión, el surco de la membrana interna y externa que las pellizca en dos mitocondrias hijas. Antes de la fisión se replican su ADN.
Muerte
Las mitocondrias al final de su vida son eliminadas por medio de la autofagia. El retículo endoplasmático celular se envuelve alrededor de la mitocondria formando una vacuola. Las vesículas del complejo de Golgi que contienen hidrolasas se unen entonces a la vacuola autofágica que luego degrada el contenido de la vacuola, la mitocondria.
Homeostasis de Ca2+
El uniportador mitocondrial en la membrana interna mueve el Ca2+ hacia la matriz que a su vez inicia la activación de las deshidrogenasas en el ciclo del ácido cítrico, las concentraciones celulares de Ca2+ esencialmente desencadenando variaciones en la actividad de la mitocondria.
Esto establece un claro vínculo entre el metabolismo de la energía y la actividad celular que implica una relación funcional con el retículo endoplasmático.
La absorción de Ca2+ es aparentemente rápida, mientras que el Ca2+ se libera de nuevo en la célula durante un período más largo, lo que indica que las mitocondrias actúan como un amortiguador. Esta función de amortiguación tiene un impacto en los potenciales terminales presinápticos y puede estar implicada en la propagación de las ondas citosólicas de Ca2+.
Neurotoxicidad y neuroprotección
La sobrecarga de las mitocondrias con Ca2+ puede estar relacionada con la apoptosis neuronal (muerte celular) a través de dos posibles resultados:
a) La disminución o pérdida del gradiente de protones de la membrana mitocondrial interna, que lleva a la terminación de la producción de ATP;
b) Pueden liberarse sustancias proactivas como el citocromo c en el protoplasma de la célula. Estas sustancias proactivas a su vez activarán caspasas que darán lugar a necrosis o apoptosis según el grado de pérdida de producción de ATP y la supervivencia de un número suficiente de mitocondrias.
Etimología u origen
proviene del griego μίτος mítos que significa "hilo" y κόνδρος kóndros que es "gránulo".
Enviar comentario o duda sobre «mitocondria»
También puedes usar el asistente de IA si prefieres una respuesta inmediata.